Los canales iónicos son proteinas de membrana ubicuas, clave para una gran variedad de procesos fisiológicos que involucran células excitables y no excitables. Mientas que el papel de los canales iónicos en las propiedades funcionales de las células excitables ha sido conocido por décadas, el estudio de su participación en la actividad de células no excitables ha sido hasta cierto punto relegado. Este es el caso de las células inmunes, como los linfocitos B y T, cuya viabilidad y regulación durante la respuesta inmune depende en gran medida en la actividad de los canales iónicos. En este artículo describimos de manera general la importancia de los canales iónicos para la respuesta inmune, las técnicas experimentales utilizadas para su estudio, los procesos celulares involucrados, y cómo se ven alterados en escenarios patológicos como la inmunodeficiencia adquirida por desnutrición.
Introducción
Los canales iónicos han sido ampliamente estudiados en una gran diversidad de tipos celulares. Su estudio inició de manera natural al tratar de dilucidar las propiedades eléctricas de las células excitables, o sea, aquellas capaces de generar actividad eléctrica en forma de potenciales de acción. Actualmente se tienen bien identificados los canales iónicos que participan en la generación de potenciales de acción en células como las de los músculos esquelético, cardiaco y liso; asimismo, de las células nerviosas y de las glándulas liberadoras de hormonas, por citar algunos ejemplos. Producto de estos estudios quedó claro que los potenciales de acción de las células excitables son producto de la apertura de canales iónicos localizados en la membrana celular con características únicas como, permanecer cerrados y abrirse sólo ante cambios de voltaje favorables en la membrana celular generando así corrientes iónicas que disparan los potenciales de acción. A este tipo de canales se les denominó voltaje dependientes. Además, se encontró que poseen una gran selectividad iónica, que en otras palabras, significa que los canales permiten el paso de sólo una especie iónica, excluyendo a las demás. Por ejemplo, los canales de sodio (Na+) y de potasio (K+) sólo permiten el paso a través de ellos de los iones Na+ y K+ respectivamente (Hille, 2001).
Otro producto importante de estos estudios fue el hallazgo de los canales iónicos que no se abren por voltaje, sino por ligando; esto es, los canales cuya apertura es promovida por la unión selectiva de una molécula o ion, por lo cual se les conoce como canales dependientes de ligando. Ahora sabemos que el papel de los canales iónicos dependientes de ligando en las células excitables es la de modular la actividad eléctrica de las células. La presencia de este tipo de canales amplió el estudio de los canales iónicos a otros tipos celulares, ya no sólo las células excitables, pensando que los canales iónicos dependientes de ligando podrían disparar y modular la respuesta celular en las células no excitables. Estudiando una gran diversidad celular se concluyó que los canales iónicos de la membrana celular son ubicuos y desempeñan un papel importante en la función y/o en el mantenimiento de su viabilidad celular (Hille, 2001).
Con estos antecedentes inició el estudio de los canales iónicos en las células no excitables como las del sistema inmune, y en particular, en los linfocitos T y linfocitos B, que tienen un rol clave en la respuesta inmune. Los linfocitos han sido ampliamente estudiados por diferentes disciplinas como la inmunología, bioquímica, biología molecular, etc., lo que ha permitido obtener un conocimiento importante del sistema inmune, destacando la descripción detallada de su activación por la presencia del antígeno y las cascadas moleculares que se disparan para iniciar y modular la respuesta inmune (Abbas et al., 2021). El estudio biofísico de los canales iónicos de los linfocitos T y B llegó tarde con respecto al resto de las disciplinas que tradicionalmente estudian la inmunología, y tal vez por esta razón no se le ha dado la importancia debida al papel que juegan los canales iónicos en la respuesta inmune (Lewis and Cahalan, 1995). En este artículo se pretende dar una visión general de la importancia de los canales iónicos en la respuesta inmune, describiendo en particular su acción en los linfocitos T. Resultados equivalentes han sido reportados para el caso de los linfocitos B.
Técnicas electrofisiológicas para el estudio de los canales iónicos.
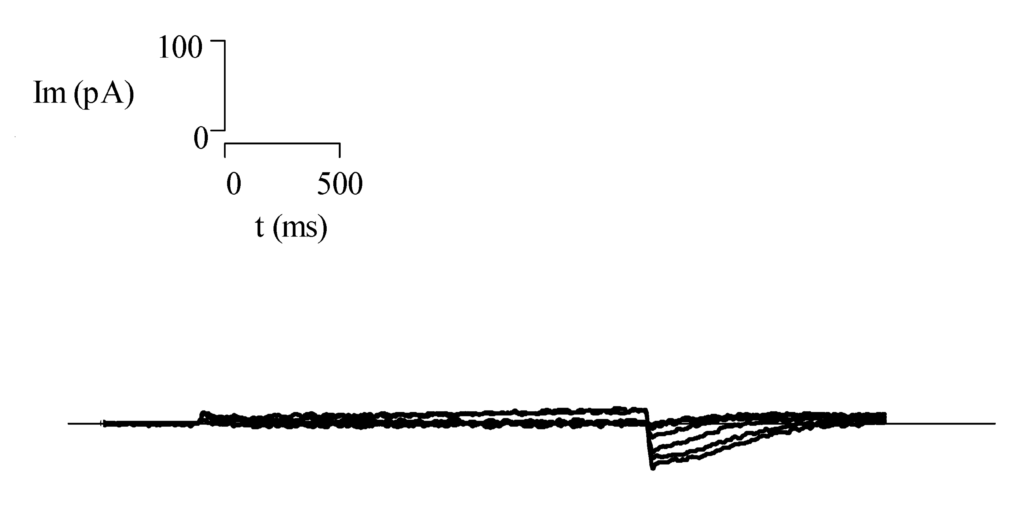
Antes de iniciar la descripción de los canales iónicos en los linfocitos T y su papel en la respuesta inmune, conviene describir brevemente una de las técnicas electrofisiológicas ampliamente utilizada para el estudio del comportamiento de los canales iónicos. Denominada Fijación del Voltaje, la técnica consiste en mantener el potencial de membrana en valores seleccionados por el usuario y por el tiempo que desee (Hille, 2001). Existen dos variedades de la técnica de fijación de voltaje, denominadas de célula completa y de registro de canal unitario. En la variedad de fijación del voltaje de célula completa (Fig. 1) se cambia el voltaje de la membrana celular para inducir la apertura de los canales iónicos de la membrana celular. Como resultado, se obtienen corrientes macroscópicas en el rango de varios cientos de picoamperes a nanoamperes (Fig 1).
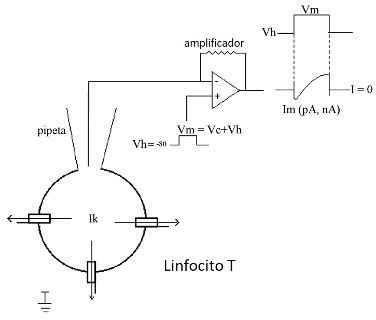
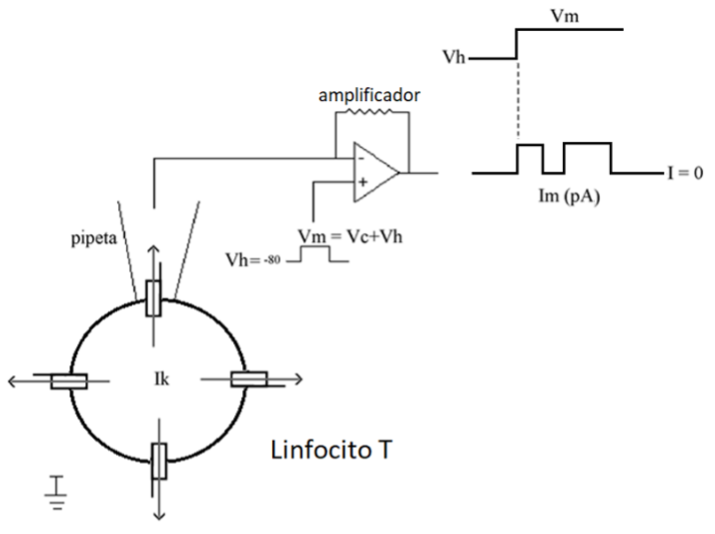
Un ejemplo de la variedad de registro de canal unitario se muestra en la Fig. 2. En esta configuración, la pipeta de registro se coloca en la superficie de la membrana celular atrapando un canal iónico. En este caso no se rompe la membrana celular y es posible medir la corriente iónica de un solo canal iónico (corriente unitaria), cuya magnitud depende del tipo de canal y fluctúa entre cientos de femtoamperes hasta varios picoamperes (Hille, 2001)
Las técnicas anteriores han sido fundamentales para el estudio de los canales iónicos en los linfocitos T. A manera de ejemplo, en la Fig. 3 se muestran registros de corrientes de K+ obtenidas en el Laboratorio de Biofísica e Ingeniería de Tejidos de la Universidad Autónoma Metropolitana, Unidad Iztapalapa. En la Fig. 3a se muestra una familia de trazos de la corriente de K+ obtenidas de linfocitos T de sangre periférica de rata Wistar. Para inducir las corrientes se aplicaron pulsos de fijación del voltaje (Vc) de diferente magnitud a partir de un potencial de mantenimiento (Vh) al cual están cerrados los canales. Cada trazo de corriente de K+ superpuesta corresponde a la inducida por un pulso rectangular de voltaje (como se expuso en la Fig. 1) como resultado de la apertura de los canales de K+. Al incrementar la amplitud de los pulsos de voltaje, la corriente de K+ aumenta, como se observa en la Fig. 3b, en donde se muestra el valor máximo de cada trazo corriente de K+ en función del potencial de membrana que indujo dicha corriente. Podemos observar que el incremento de la corriente de K+ con respecto al cambio en el potencial de membrana no es lineal.
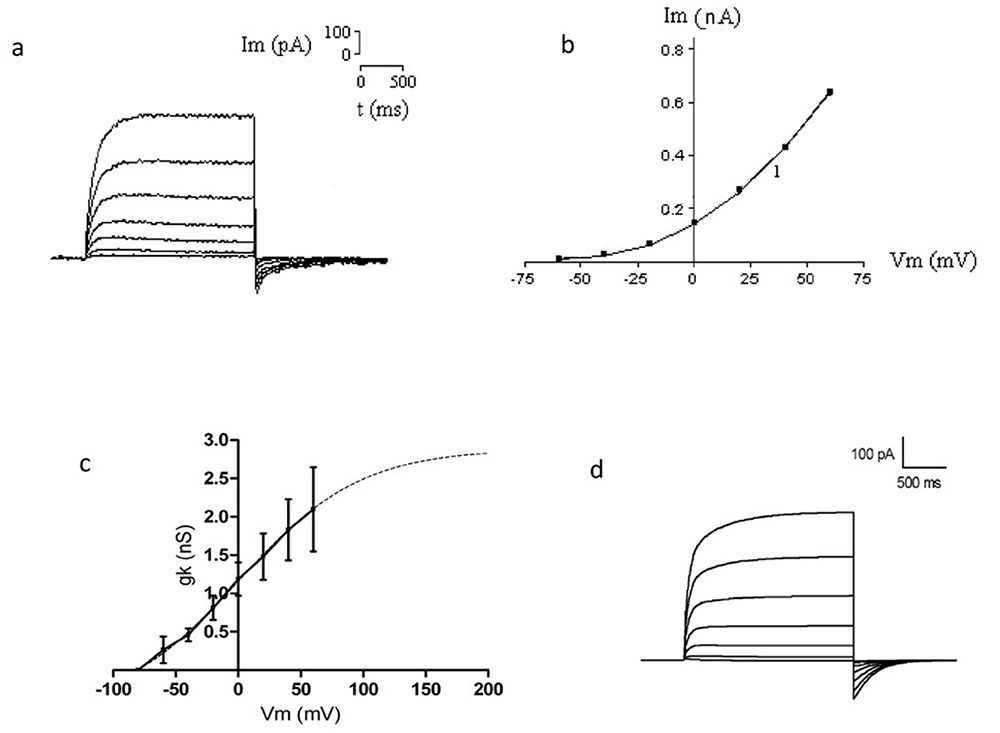
En la Fig 3c se muestra la curva de conductancia al K+ (gK+) con respecto al potencial de membrana (gK+ vs Vm). La conductancia se obtiene a partir de los datos de la curva corriente de K+ vs Vm (Fig. 3b). Con los datos de las Figs. 3a-c es posible obtener un modelo teórico de la apertura de los canales de K+ que reproduce el curso temporal de la corriente de K+ y la corriente máxima que se obtiene para cada pulso de voltaje (Fig. 3d), esto es:
I_{K^+}=gK^+(V_m,t)(V_m-V_K)En la ecuación anterior, la conductancia al K+ (gK+) es función del voltaje de membrana Vm y del tiempo t y Vk indica el potencial de equilibrio del K+. La metodología para obtener gK+(Vm,t) a partir de los datos experimentales es estándar y se realiza de rutina por los biofísicos experimentales que estudian a una determinada población de canales iónicos (Hille, 2001).
La metodología de fijación del voltaje para la obtención de la corriente iónica que ha sido descrita brevemente, que nos lleva a la construcción de gráficas como las mostradas en la Fig. 3 y la generación de modelos cuantitativos a partir de los datos experimentales, ha sido empleada también para el estudio de las diversas poblaciones de canales iónicos presentes en los linfocitos T, además de los canales de K+.
Canales iónicos en los linfocitos T no activados (vírgenes).
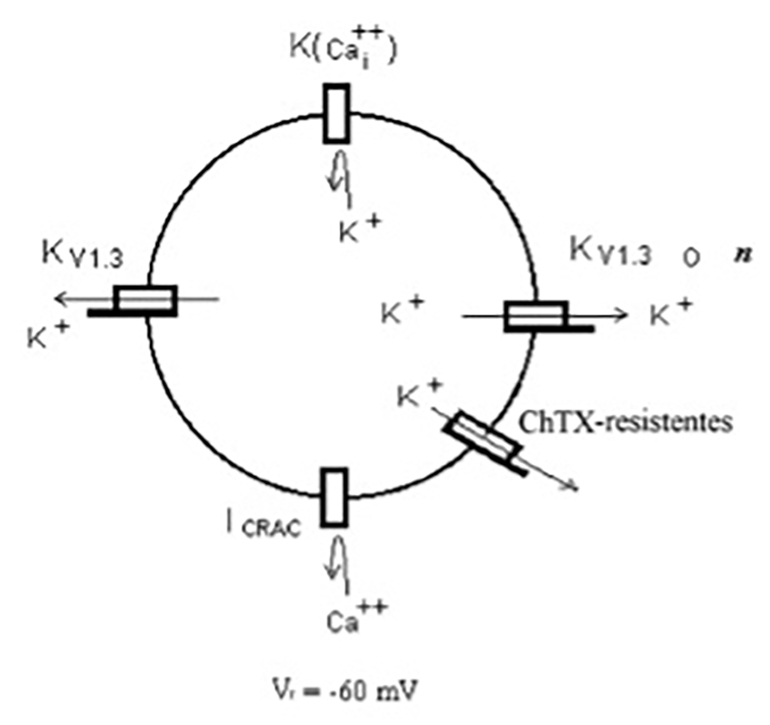
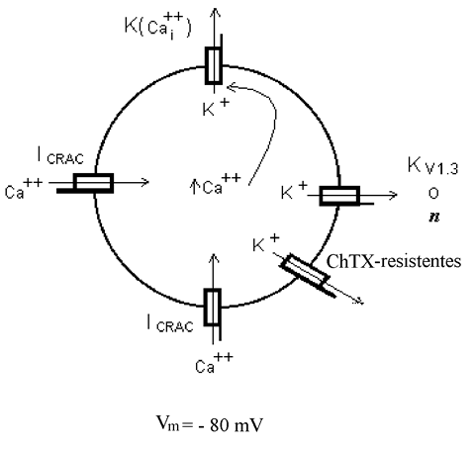
Los linfocitos T vírgenes son aquellos que no han sido activados por la presentación de algún antígeno (Zweifach and Lewis, 1993). Estos presentan varios tipos de canales iónicos en la membrana celular entre los que destacan los canales de K+ denominados KV1.3 como se ilustra en el esquema de la Fig. 4. Asimismo, en menor proporción, la membrana celular presenta otra población de canales de K+ denominada resistente a Charibdotoxina (ChTX) ya que, a diferencia de los canales KV1.3, no son bloqueados por el fármaco ChTX. Los canales KV1.3 y ChTX-resistentes se encuentran abiertos permitiendo el paso de K+ a través de ellos, lo que origina un voltaje de membrana negativo denominado potencial de reposo (Vr) cuyo valor promedio es de -60 mV (Lewis and Cahalan, 1995).
Los linfocitos T presentan además canales de K+ activados por calcio (Ca++), que comúnmente se designan como K(Ca++). Como se muestra en la Fig. 4, los canales de K(Ca++) se encuentran normalmente cerrados impidiendo el paso de K+. Estos canales se abren por ligando al unir iones Ca++ del lado citoplasmático del canal. Ya que la concentración de Ca++ intracelular [Cai++] es muy baja, alrededor de 10-7 Molar, la probabilidad de unión del Cai++ al canal para su apertura es muy baja y como consecuencia el canal permanece cerrado. Otro canal presente en la membrana celular del linfocito T es el canal de Ca++ denominado CRAC, que también permanece cerrado impidiendo el paso de Ca++ a través de él. La apertura del canal CRAC es dependiente de dos factores: el Vm y un ligando. En cuanto al Vm, el canal se encuentra cerrado al potencial de membrana de reposo de -60 mV, valor común en un linfocito T que no ha sido activado por la presencia de algún antígeno, y se abre cuando el linfocito T se hiperpolariza. El ligando, denominado STIM1, es una molécula mensajera que se produce cuando los depósitos intracelulares de [Ca++]i se agotan (Zweifach and Lewis, 1993).
Eventos intracelulares durante la fase inicial de la activación del linfocito T
Un linfocito T virgen inicia una cascada de eventos intracelulares cuando une de manera específica su antígeno como se ilustra en la Fig. 5. Los linfocitos T exhiben en su superficie un receptor específico (RTC) para su antígeno (Ag), que es presentado por otras células del sistema inmune denominadas células presentadoras de antígeno. Los antígenos son trozos moleculares de agentes infecciosos. ¿Cómo es que estos trozos moleculares o antígenos son presentados a los linfocitos T? Los eventos son los siguientes: cuando un agente infeccioso ingresa al organismo, células del sistema inmune inespecífico, como los macrófagos, fagocitan al agente invasor, destruyendo al microrganismo en su interior. Trozos moleculares del agente patógeno (antígeno) ahora son presentados por los macrófagos a los linfocitos T para iniciar su activación. Una vez que el Ag se une al RTC se activa la enzima fosfolipasa C-ϒ1, que actúa sobre el sustrato 4,5 bifosfato de fosfatidilinositol (FIB) para descomponerlo en DG e IP3. Por otro lado, los linfocitos T tienen un retículo endoplásmico (RE) que actúa como un almacén intracelular de Ca++ debido a las bombas de Ca++ en su membrana que permiten la entrada y acumulación del Ca++ en su interior. El IP3 que se formó por la unión del Ag al RTC, actúa sobre el RE abriendo los canales de Ca++ presentes en su membrana. Como resultado, el Ca++ saliente del RE hacia el citoplasma incrementa la concentración de Ca++ intracelular [Cai++.]. Se sabe que este incremento inicial del Cai++ es crucial para el inicio de la activación del linfocito T; sin embargo, la activación requiere de un incremento sostenido del Cai++, lo que hace a los depósitos intracelulares de Cai++ insuficientes al agotarse rápidamente (Fomina et al., 2000; Zweifach and Lewis, 1993).
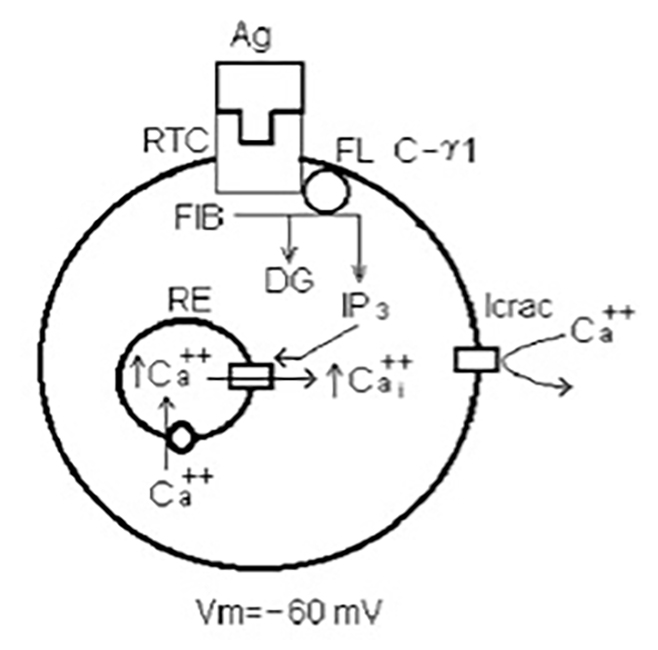
El incremento sostenido del Cai++ indispensable para la activación del linfocito T es debido a la entrada de Ca++extracelular por la apertura de los canales CRAC de Ca++. El mecanismo para la apertura de los canales CRAC es el siguiente (Fig. 6): El incremento inicial de la [Cai++], que como ya se comentó, proviene del retículo endoplásmico (RE), abre el canal de K(Ca++) que produce un potencial de membrana más negativo que el potencial de reposo Vr, adquiriendo un valor de -80 mV. Esta hiperpolarización es indispensable para la apertura del canal CRAC pero insuficiente. La apertura del canal CRAC se da finalmente por la acción concertada de la hiperpolarización y el mensajero intracelular STIM1 que se produjo al agotarse el RE de Ca++. El linfocito T se activa apropiadamente al mantener elevada la [Cai++] de manera sostenida, producto de la entrada de Ca++ vía los canales de Ca++ CRAC abiertos, produciendo la corriente de Ca++ ICRAC (Fomina et al., 2000; Zweifach and Lewis, 1993).
Como podemos observar, los canales iónicos son fundamentales en la activación de los linfocitos T, etapa inicial de la respuesta inmune ante la presencia de antígenos. La alteración de los mecanismos descritos puede redundar en una activación pobre o nula de los linfocitos T. Como un ejemplo importante de falla en el proceso de activación de los linfocitos T se encuentra la inmunodeficiencia adquirida por la desnutrición grave.
Inmunodeficiencia Adquirida por la Desnutrición Grave y los Canales iónicos
Desde antes de la medicina moderna se sabía que las hambrunas en las poblaciones humanas las hacían más susceptible a padecer infecciones mortales (Beatty y Dowdle, 1978; Blair et al., 2002). Actualmente, diversas disciplinas científicas han puesto de manifiesto una diversidad de factores que alteran e inhiben la respuesta inmune durante la desnutrición grave (Chandra, 1999); sin embargo, uno que no se había planteado, es si la desnutrición grave podría afectar a los canales iónicos, que como se describió, son fundamentales para la activación del linfocito T. Es natural proponer que la desnutrición grave los afecte ya que la pobre ingesta de proteínas y calorías afectaría la producción de las proteínas que fungen como canales iónicos en los linfocitos T. En particular, como se comentó, los canales de K+ son claves para hiperpolarizar la membrana celular y hacer posible la apertura de los canales de Ca++ CRAC para el incremento sostenido de Cai++ que sucede durante la activación. Una población de canales de K+ disminuida reduciría la conductancia al K+, imposibilitando la hiperpolarización y la activación del linfocito T. Este sería otro factor más que se sumaría a los ya descritos en la desnutrición grave. En la Fig. 7 se muestran los trazos de corriente de K+ de un linfocito T obtenido de ratas Wistar con desnutrición grave. En ella podemos observar que las corrientes de K+ están muy disminuidas al compararlas con el caso control mostrado en la Fig 3a.
Con este resultado, se muestra que otro factor importante en la respuesta inmune deficiente durante la desnutrición grave, es la disminución en la conductancia al K+, que imposibilitaría la hiperpolarización necesaria para la activación de los canales CRAC que mantienen elevada la [Cai++] indispensable para la activación del linfocito T. El resultado es la incapacidad de respuesta del sistema inmune específico ante agentes infecciosos, aumentando la probabilidad de muerte por infección.
Conclusión
Los canales iónicos son fundamentales para la activación de los linfocitos T (Cahalan y Chandy, 2009). La activación de los linfocitos T es la etapa de arranque de la respuesta del sistema inmune a la presencia de antígenos, los cuales son componentes moleculares de agentes patógenos de tipo infeccioso. Esta etapa es crítica para una respuesta inmune adecuada, la cual culmina con la destrucción de agentes infecciosos. Como se comentó, en esta etapa los canales iónicos juegan un rol importante y la alteración de su función puede afectar la activación linfocitaria impidiendo o limitando la respuesta inmune. Existen en la actualidad una diversidad de agentes farmacológicos conocidos como inmunomoduladores que se emplean en la clínica para incrementar la respuesta inmune y para disminuir la respuesta inmune como en le caso de las enfermedades autoinmunes. Por otro lado, la biofísica de canales iónicos ha mostrado cómo el comportamiento de diferentes poblaciones de canales iónicos puede cambiar por la acción de diferentes agentes farmacológicos. Estos hallazgos tienen el potencial de ser empleados en la clínica para modular la actividad de los canales iónicos y a su vez, la respuesta inmune. En otras palabras, el conocimiento de los canales iónicos en la respuesta inmune abre la posibilidad de emplear otra batería de fármacos que afecten a los canales iónicos de tal manera que harían posible modularla, aumentando su acción en el caso de infecciones graves o tratamiento del cáncer (Ginefra et al., 2021) y limitando su acción en el caso de las enfermedades autoinmunes.
Referencias
Abbas, A., Lichtman, A., Pillai, S. Cellular and Molecular Immunology, Elsevier, Philadelphia, 2022.
Beatty, D. y Dowdle, E. The effects of Kwashiorko serum on lymphocyte transformation “in vitro”. Clin. Exp. Inmunol. [32], pp. 134-143, 1978.
Blair, S., Carmona, J. y Correa, A. Malaria en niños: relaciones entre nutrición e inmunidad. Rev. Panam. Salud Pública [11], pp. 5-14, 2002.
Cahalan, M. and Chandy, K., The functional network of ion channels in T lymphocytes. Immunol. Rev. [231(1)], pp. 59–87, 2009
Chandra, R. Nutrition and the Immune System: an introduction. Am. J. Clin. Nutr. [66(2)], pp. 460S-463S, 1997.
Fomina, A., Fanger, C., Kozak, J. y Cahalan, M. Single channel properties and regulated expression of Ca2+ release-activated Ca2+ (CRAC) channels in human T cells. J. of Cell Biol. [150], pp. 1435-1444, 2000.
Ginefra, P., Carrasco, H., Spagna, M., Zecchillo, A., y Vannini, N. Ionic Regulation of T-Cell Function and Anti-Tumour Immunity. Int. J. Mol. Sci.[22], pp. 13668, 2021.
Hille, B. Ion Channels of Excitable Membranes, Sinauer Associates Inc. Publishers Sunderland, Massachusetts, 2001.
Lewis, R. y Cahalan, M., Potassium and calcium channels in lymphocytes. Annu. Rev. Immunol. [13], pp. 623-653, 1995.
Zweifach, A. y Lewis, R. Mitogen-regulated Ca2+ current of T lymphocytes is activated by depletion of intracellular Ca2+ stores. Proc. Natl. Acad. Sci. USA. [90], pp. 6295-6299, 1993.